
下载中心
download center
为什么水产品不会携带2019新型冠状病毒
中国是世界水产养殖大国。2018年,全国水产养殖面积7189.52千公顷,养殖产量4991.06万吨,出口量432.2万吨、出口额223.26亿美元,渔业从业人员1325.72万人,渔民人均纯收入19885.00元(2019中国渔业统计年鉴)。水产养殖在保障我国优质蛋白供给、调整农业结构和振兴乡村经济等方面发挥着重要作用。
2019年12月,湖北武汉暴发了由2019新型冠状病毒(2019 novel coronavirus,2019-ncov)感染引起的新冠状病毒肺炎(novel coronavirus pneumonia,ncp)[1,2]。研究人员对早期确诊病例的调查发现,武汉华南海鲜批发市场与此次肺炎疫情有密切的关系[3]。武汉华南海鲜批发市场名义上是海鲜市场,但实际上为综合农贸市场,市场的西区主要从事野生动物交易。肺炎疫情发生后,研究人员对该市场采集的585份环境样本进行检测,33份样品呈2019-ncov阳性,其中93.9%(31/33)阳性样本分布在市场的西区,即表明2019-ncov来源于武汉华南海鲜批发市场销售的野生动物。但是,由于武汉华南海鲜批发市场主要的销售对象为水产品,水产品安全问题不可避免的成为了人们关注的焦点。从目前发展形势看,此次新冠状病毒肺炎疫情已对我国水产养殖业造成了一定程度的影响。如何让大众正确认识2019-ncov与水产品的关系,促进水产品生产,让市场销售尽早回归正常,成为当前水产养殖业必须解决的严峻问题。本文以病毒对宿主选择的特点为切入点,浅析2019-ncov与水产品间的关系,旨在为消除消费者对水产品顾虑、保障水产品有效供给、打赢疫情防控阻击战提供一定的科技支撑。
冠状病毒(coronaviruses,covs)属于巢状病毒目(order:nidovirales)、冠状病毒科(family:coronaviridae)、正冠状病毒亚科(subfamily:orthocoronavirinae),该亚科包括四个属:α-冠状病毒(genus:alphacoronavirus),β-冠状病毒(genus:betacoronavirus),γ-冠状病毒(genus:deltacoronavirus)和δ-冠状病毒(genus:gammacoronavirus)[4]。冠状病毒为不分段的单股正链rna[ssrna( )]病毒,完整的病毒颗粒呈圆形或椭圆形,直径约60~220 nm之间,大小可达26-32 kb,有囊膜,外周有冠状排列的刺突蛋白(spike蛋白,s蛋白),因此被命名为冠状病毒[5] (图1)。
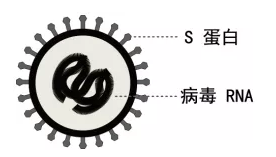
冠状病毒可以感染人类、牲畜、禽类、蝙蝠、小鼠和许多其他野生动物,引起宿主呼吸系统、胃肠道、肝脏和中枢神经系统疾病,其中β-冠状病毒对人类的危害最为严重[6,7]。2003年中国暴发的重症急性呼吸综合征 (severe acute respiratory syndromes,sars)和2012年沙特阿拉伯暴发的中东呼吸综合征(middle east respiratory syndrome coronavirus,mers),其病原分别为重症急性呼吸综合征冠状病毒(severe acute respiratory syndromes coronavirus,sars-cov)和中东呼吸征冠状病毒(middle east respiratory syndromes coronavirus,mers-cov),两种病毒都属于β-冠状病毒[8,9]。2019年12月,湖北省武汉市暴发的新冠状病毒肺炎,其病原2019-ncov也属于β-冠状病毒[2]。
冠状病毒具有相对严格的宿主特异性,只能感染天然宿主和亲缘关系及其相近宿主[10],其中β-冠状病毒主要感染人和其他多种哺乳类动物[11] 。
sars 疫情暴发后,研究人员从多处市场销售的果子狸样品体内分离出与sars-cov非常相似的sars样冠状病毒,将果子狸来源的sars样冠状病毒与人sars-cov进行基因比较,证实了sars-cov来源于市场内销售的果子狸[12-13]。随后研究发现,仅能从农贸市场和饲养场中的果子狸体内能检出sars-cov,未能从野外捕获的果子狸体内检出,来源于不同果子狸的sars-cov基因有很高的非同义/同义替换率,提示果子狸可能只是sars-cov从动物到人传播的中间宿主[13-15]。2005年,通过对408只蝙蝠的血清、咽拭子和肛拭子样品进行检测,成功获得一株与sars-cov基因组高度相似的sl-cov,病毒呈现丰富的遗传多样性,证实了中华菊头蝠为sars的自然宿主[16]。
2012年 mers 疫情在沙特阿拉伯暴发后,将获得的mers-cov基因与不同来源蝙蝠体内的冠状病毒基因比较发现,mers-cov与扁颅蝠冠状病毒 hku4 和伏翼蝙蝠冠状病毒 hku5 亲缘关系最为接近[17,18] 。然而,从生态位的角度考虑,蝙蝠与人类的接触几率少,且从蝙蝠中检出的阳性率较非常低,因此研究人员推测,在蝙蝠和人之间,可能存在一种中间宿主,作为病毒的增殖载体,使其更易于传播给人类[19]。alagaili等[20]在沙特全国范围内采集的200多头单峰驼血液样本中,有74%的样本血清中mers-cov抗体呈阳性。2014年,对一名感染mers死亡男子和其饲养的单峰骆驼体内分离的病毒进行比对,发现不同来源的病毒基因组是一致的,表明死者因为与患病单峰骆驼密切接触而被传染,证实了单峰骆驼是mers-cov的中间宿主[21]。
2019年新冠状病毒肺炎暴发后,石正丽研究员团队[1]通过基因组序列比较发现,2019-ncov与来源于中华菊头蝠样本的一株冠状病毒(ratg13)的基因相似,两种病毒序列一致性高达96.2%。上述研究结果表明,与sars-cov和mers-cov一样,2019-ncov的自然宿主也是蝙蝠,但其中间宿主是何种动物,目前尚未可知。
sars-cov是从蝙蝠传染到果子狸,再由果子狸传染给人,后通过和被感染者密切接触而在人与人之间传播[22];mers-cov是起源于蝙蝠体内的冠状病毒,经单峰骆驼传给人[21]。人、蝙蝠、果子狸、骆驼都属于哺乳类,目前已报道2019-ncov的自然宿主也是蝙蝠,根据β-冠状病毒具有宿主特异性的特点,2019-ncov的中间宿主很有可能是一种哺乳动物(图2)。基于上述分析,初步排除了水产动物成为2019-ncov宿主的可能。

病毒的感染首先依赖于吸附蛋白对易感细胞表面受体的吸附,吸附是启动病毒感染的第一步,也是决定病毒感染能否成功的关键环节。冠状病毒s蛋白细胞受体的鉴别对揭示病毒入侵、跨物种传播等机制有重要意义[1]。li等[23]发现从非洲绿猴细胞株vero e6细胞筛选出的血管紧张素转化酶2(angiotensin-converting enzyme 2,ace2)可与sars-cov的s蛋白(纤突蛋白)产生特异性结合;sars-cov能在转染了ace2的人肾细胞株293t细胞(原始的293t细胞对sars-cov不敏感)中有效的增殖,而抗ace2的抗体可与阻止病毒的增殖,证明了ace2是sars-cov入侵宿主细胞的关键受体蛋白。随后,li等[24]将人、小鼠及大鼠的ace2基因分别转染到293t细胞中,再用含有sars-cov s基因的siv假病毒对转染后的细胞进行感染,结果显示,小鼠和大鼠ace2转染细胞的病毒进入效率明显低于人ace2转染细胞的病毒进入效率,将人ace2转染小鼠3t3细胞则可使病毒的进入效率明显增加,表明不同物种间ace2的差异对病毒入侵具有一定影响。
2019-ncov s蛋白的细胞受体也是ace2,2019-ncov对细胞的入侵依赖于细胞受体ace2[1]。利用megalign 软件对来源于人、小鼠、大鼠、蝙蝠、果子狸、斑马鱼的aec2氨基酸进行遗传距离分析。结果显示,哺乳动物间ace2氨基酸的相似性,在77.7%~91.3%之间,而斑马鱼与其他哺乳动物的ace氨基酸的相似性,在74.7%~78%之间(图3)。基于不同动物ace2氨基酸序列,采用neighbor-joining 法构建的进化分析结果显示,与哺乳动物相比,斑马鱼被单独为一簇(图4)。上述结果表明,鱼和哺乳动物间有较远的亲缘关系,且鱼与哺乳动物的ace2氨基酸序列差异较大,推测其蛋白的结构也一定存在较大差异,从而影响2019-ncov s蛋白与鱼类细胞的结合。因此,鱼类ace2很难介导2019-ncov进入鱼体细胞,2019-ncov不能利用鱼体细胞进行增殖,从而进一步证明了水产动物不能成为2019-ncov的宿主。
注:对角线以上为相似性百分比,对角线以下为散度百分比。

danio rerio ace2:斑马鱼ace2,genbank登录号nm_001007297.1;homo sapiens ace2:人ace2,genbank登录号nm_001371415.1;mus musculus ace2:小鼠ace2,genbank登录号nm_001130513.1;rattus norvegicus ace2:大鼠ace2,genbank登录号nm_001012006.1;rhinolophus ferrumequinum ace2:马铁菊头蝠ace2,genbank登录号ab297479.1;rousettus leschenaultii ace2:棕果蝠ace2,genbank登录号gu253336.1;paguma larvata ace2:果子狸ace2,genbank登录号ay881174.1
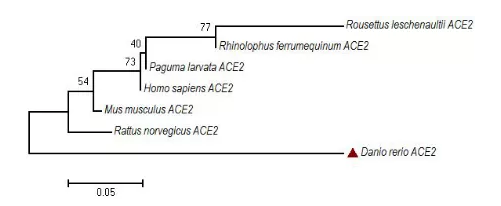
danio rerio ace2:斑马鱼ace2;homo sapiens ace2:人ace2;mus musculus ace2:小鼠;rattus norvegicus ace2:大鼠ace2;rhinolophus ferrumequinum ace2:马铁菊头蝠ace2;rousettus leschenaultii ace2:棕果蝠ace2;paguma larvata ace2:果子狸ace2
冠状病毒有较为严格的宿主特异性,病毒能否与宿主细胞受体发生特异性结合是病毒选择宿主的关键环节。目前,已有较多关于2019-ncov宿主的报道,包括蝙蝠、水貂(未确定)、穿山甲(未确定)等哺乳动物。为了解除大众对水产动物是否携带2019-ncov的疑惑,本文以sars-cov和mers-cov为例,阐述了两种烈性β-冠状病毒的溯源过程,明确了β-冠状病毒主要以哺乳动物作为宿主;根据水产动物和哺乳动物的ace2氨基酸序列差异性,分析出2019-ncov不能在水产动物体内增殖。基于以上结果,排除了水产动物为2019-ncov宿主的可能性。
此外,在中国营养学会编著的《中国居民膳食指南》中指出,鱼富含优质蛋白质、脂类、脂溶性维生素、b族维生素和矿物质等;与畜禽类和蛋类相比,鱼类脂肪含量相对较低,且含有较多的不饱和脂肪酸,对预防心脑血管和血脂异常疾病等具有重要作用。根据指南的推荐,大家每日水产品的摄入量应该达到40克以上,而我国每日人均摄入量仅为30克左右。从目前病例统计数据看,新冠状病毒肺炎病情危重的患者多为高龄人员,部分患者有肺部或心血管基础疾病。因此,新冠状病毒肺炎疫情期间,吃水产品不仅安全,适当增加每日水产品的摄入量还将有效的改善心血管疾病,有助于新冠状病毒肺炎的预防。
综上所述,总结为以下三点:(1)鱼的病毒不能在人体内复制,人的病毒也不能在鱼体内增殖;(2)没有发现任何水产养殖动物以及水生动物病毒与2019新型冠状病毒有任何直接联系;(3)水产品是健康的,食用水产品是安全的,水产品不会传染新冠状病毒肺炎。
1. zhou p, yang xl, wang xg, et al. discovery of a novel coronavirus associated with the recent pneumonia outbreak in humans and its potential bat originbio. rxiv, 2020, doi: https://doi.org/10.1101/2020.01.22.914952.
2.wu f, zhao s, yu b, et al. a new coronavirus associated with human respiratory disease in china. nature, 2020, https://doi.org/10.1038/s41586-020-2008-3.
3.huang c, wang y, li x, et al. clinical features of patients infected with 2019 novel coronavirus in wuhan, china. lancet. 2020 jan 24. pii: s0140-6736(20) 30183-5.
4.international committee on taxonomy of viruses, taxonomy history: cornidovirineae[eb/ol].https://talk.ictvonline.org/taxonomy/p/taxonomy history? taxnode_id=20186105 (accessed on 2 january 2020).
5.almeida jd, berry dm, cunningham ch. coronaviruses[j]. nature, 1968, 220: 650-659.
6.corman vm, muth d, niemeyer d, et al. hosts and sources of endemic human coronaviruses[j]. advances in virus research, 2018, 100:163-188.
7.cui j, li f, shi zl. origin and evolution of pathogenic coronaviruses [j]. nat rev microbiol, 2019, 17(3):181-192.
8.li f, li wh, farzan m, et al. structure of sars coronavirus spike receptor-binding domain complexed with receptor[j]. science, 2005, 309(5742): 1864-1868.
9.de groot r , baker s , baric r , et al. middle east respiratory syndrome coronavirus (mers-cov): announcement of the coronavirus study group[j]. journal of virology, 2013, 87(14):7790-7792.
10.enserink, m. infectious diseases: calling all coronavirologists[j]. science, 2003, 300(5618):413-414.
11.熊成龙, 蒋露芳, 姜庆五. β-冠状病毒引起人类疾病的流行与控制[j]. 上海预防医学, 2020, 32(1): 1-12.
12.guan y, zheng bj, he yq, et al. isolation and characterization of viruses related to the sars coronavirus from animals in southern china [j]. science, 2003, 302: 276-278.
13.song hd, tu cc, zhang gw, et al. cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human [j]. proc natl acad sci u sa, 2005, 102(7): 2430-2435.
14.guan, y, peiris, jsm, zheng, b, et al. molecular epidemiology of the novel coronavirus that causes severe acute respiratory syndrome[j]. lancet, 2004, 363(9403): 99-104.
15.kan b, wang m, jing h, et al. molecular evolution analysis and geographic investigation of severe acute respiratory syndrome coronavirus-like virus in palm civets at an animal market and on farms [j]. j virol, 2005, 79(18): 11892-11900.
16. li wd, shi zl, yu m, et al. bats are natural reservoirs of sars-like coronaviruses[j]. science, 2005, 310(5748): 676-679.
17. van boheemen s , de graaf m , lauber c , et al. genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans[j]. mbio, 2012, 3(6): e00473-12-e00473-12.
19. memish za, mishra n, olival kj. middle east respiratory syndrome coronavirus in bats, saudi arabia. emerg infect dis, 2013, 19(11): 1819-1823.
20. alagaili an , briese t , mishra n, et al. middle east respiratory syndrome coronavirus infection in dromedary camels in saudi arabia[j]. mbio, 2014, 5(2): e00884-14.
21. drosten c , kellam p , memish za. evidence for camel-to-human transmission of mers coronavirus[j]. new england journal of medicine, 2014, 371(14): 1359-1360.
22. graham r l, baric rs. recombination,reservoirs, and the modular spike: mechanism s of coronavirus cross-species transmission [j].j virol, 2010, 84 (7): 3134-3146.
23. li wh, moore m j, vasilieva n, et al. angiotensin-converting enzyme 2 is a functional receptor for the sars coronavirus[j]. nature, 2003, 426(6965): 450-454.
24. li wh, greenough tc, morem j, et a1. efficient replication of severe acute respiratory syndrome coronavirus in mouse cells [j]. j virol, 2004, 78 (20): 11429 -11433.